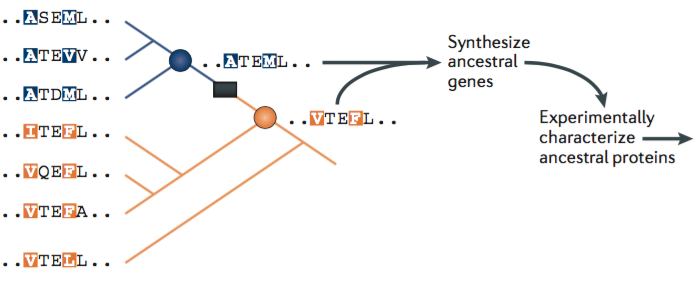
With this episode I would like to do some systematic theology and focus on the most foundational subject of theology: God. Systematic theology is theology that pursues an orderly, rational, and coherent method. There are benefits to the systematic, orderly approach, which I want to take advantage of here. But it is admittedly not characteristic of the texts of scripture, which are often disorderly, uncanny, and occasionally contradictory. The systematic approach is a convenient way to understand and analyze theological concepts, but it’s usually not the way we actually encounter these things in religious experience. I’m reminded here of Blaise Pascal’s statement: “God of Abraham, God of Isaac, God of Jacob, not of the philosophers.” There’s much to be said for that sentiment. Nevertheless the systematic approach still has significant utility for comprehension and analysis. In talking about God in this systematic way the understanding of God I will take is that of classical theism.
In what follows I just want to lay out what classical theism is. I won’t get too much into arguments or proofs for God or for classical theism. That’s another topic. But I hope that just presenting what classical theism is will show it to be a very plausible and reasonable thing to believe. Even before taking any steps to argue for it or prove it.
First some definitions. Theism is the belief in the existence of God or gods. Monotheism is the belief that there is only one God. Classical theism is the belief that God is the source of all things. In more technical terms classical theism is the belief that God is metaphysically absolute. Classical theism is a form of monotheism but it’s more theoretically developed. It takes the belief that there is only one God and analyzes what that means, the way in which there is only one God, what this one God must be like. This is what makes it systematic, theological, and philosophical.
What does it mean for God to be metaphysically absolute, the source of all things? There are two major ways for there to be only one God. They are quite different and imply very different things about God’s nature. One way is for there to be a pre-existing reality in which God exists, a reality that is independent of God and prior to God. There’s a universe that happens to have a God in it and there’s only one God. The other way, the way of classical theism, is for God to be prior to everything. There is nothing without God. All reality depends on God for its existence. We could think of these loosely as God being inside all reality versus God being outside or beyond all reality.
In classical theism all of reality derives from God and depends on God. It’s even possible for God to be the only thing that exists. But it’s actually not possible for God not to exist. This is to say that God is absolutely necessary. Nothing else is necessary in this way. Everything else is contingent. It is possible for everything else not to exist. But it is not possible for God not to exist.
Classical theism tends to be philosophical, trans-religious, and trans-scriptural, meaning that it spans many religions and the texts of many religious traditions. Throughout history classical theists have been Christian, Pagan, Jewish, Muslim, and Hindu. Obviously classical theists in each of these traditions disagree on a lot. But they tend to agree in their classical theism and in their understanding of God’s primary attributes, even if they disagree on the specific things they believe God to have done in human history. Pagan classical theists include Plotinus and Proclus. Jewish classical theists include Philo of Alexandria and Maimonides. Christian classical theists include Augustine, Pseudo-Dionysius, Anselm, and Thomas Aquinas. Muslim classical theists include Ibn Sina, and Ibd Rushd. I also think that many of the ideas of Hindu thinkers like Shankara and Ramanuja have much in common with classical theism.
What’s interesting about classical theism is that it basically starts from the premise of God’s metaphysically absolute nature and derives God’s attributes from there. These attributes often coincide with scripture, albeit not always perfectly, which is an important theological issue. But that’s also a topic for another time. The attributes of God in classical theism include the following:
Aseity
Necessity
Simplicity
Eternity
Immutability
Immateriality
Omnipotence
Omniscience
Perfect Goodness
Aseity is not a well-known term but it’s very important to the topic. The word comes from Latin “a se” meaning “from self”. Aseity is the property by which a being exists of and from itself, and not from anything else. God’s aseity means that God does not depend on anything else for his existence; not on the universe, not on anything it all.
Necessity is when something cannot fail to be the case. For example, logical truths are generally considered to be necessarily true. An example would be the proposition “If p and q, then p”. This would seem to be necessarily true. It couldn’t be otherwise. Philosophers might still debate that but it should at least be clear what we’re talking about with necessity. God’s necessity means that God cannot not exist. Understanding why that is and arguing for it is a bigger topic. But understanding the claim that God is necessary is key to understanding what classical theism is.
Simplicity means not having any parts. According to classical theism God is simple in this way. God is not composed of parts. Put another way, God is not composite. Composite is the opposite of simple. Many philosophers consider divine simplicity to be the most important concept of classical theism and hold that all of classical theism derives from it and is ultimately equivalent to it. To understand some of the motivation behind this, anything that is composite, made up of parts, has to be put together in the way that it is put together. But composition of this kind makes it dependent on whatever it is that puts it together. So it wouldn’t be the first or source of all things.
Eternity refers to what exists outside of time. Eternity, as understood in classical philosophy, is different from how the word is commonly understood. There is the notion of things being everlasting, existing within time but lasting forever, for an infinite duration. But this is different from the kind of eternity in classical theism. God’s eternity is his existence outside of time itself. Time, in fact, would be one of the things created by God. We can imagine God looking at the passage of time as we look at the passage of time for characters in a book. For the characters in a story, if they were real, they would experience time sequentially. But for us as readers we can look at the story as a whole, all at once, because we are outside of the time of that story. Like the characters in that story, we experience our time sequentially. The past is behind us. The future is ahead of us. Only the present is before us. But for God it is all present and equally before him.
Immutability is the impossibility of changing. There’s definitely a relation here to eternity. God could hardly change across time since he exists outside of time itself. This brings up an interesting question about whether God, being immutable, will seem the same to us at all times. Not necessarily. Even if God doesn’t change, we do. For example, God is perfectly good and that doesn’t change. But our morality varies significantly. The way we perceive God will vary significantly depending on whether our conduct is mostly moral versus mostly immoral.
Immateriality, as the term suggests, is the quality of not being material. Even without a technical definition I think we all have a good intuition what materiality is. In fact, it’s more difficult to think of anything that isn’t material. It’s the material that makes up our immediate experience. Matter is the stuff that, when you kick it, it kicks back. Material things exist in time and space. If we refer to more modern chemistry and physics, matter is composed of particles, waves, and fields. Particles like protons, neutrons, and electrons have mass, particles like photons do not. But they’re all material. Material things interact with each other. They exchange momentum; they attract or repel each other through electric change. Photons induce chemical reactions. But God, being immaterial is not like any of these things.
How could any thing be immaterial? This was a question that Augustine had. He was finally able to conceptualize immaterial entities by way of Platonist and Neo-Platonist philosophy, which have a lot to say about immaterial forms. Today we most commonly come across immateriality in the form of abstract, mathematical, and logical objects. The philosopher Phillip Cary uses the example of the Pythagorean theorem. The Pythagorean theorem is not something that exists in space and time. It’s eternal, necessary, and omnipresent. It didn’t ever start being true and it will never stop being true. It cannot not be true. And it’s true everywhere. It’s not made up of particles, waves, or fields. It’s not something you handle or that kicks back. That gives an idea of what an immaterial thing can be like.
God is not an abstract, mathematical, or logical object. But he is immaterial in classical theism. He’s more like an abstract, mathematical, or logical object than he is like an electron, proton, or magnetic field.
Omnipotence is the quality of having unlimited power. This is very related to God’s nature as metaphysically absolute, the source of all things. All things come from God and are the way they are because of God. There is no other source for all that is and no other power in serious competition with God. God is able to do anything that it is possible to do. What kind of constraints does that condition impose? What would be impossible for God? Contradiction certainly. Even God cannot make something to be the case and not be the case. You’ve probably heard the question, often asked in jest, “Could God make a stone so heavy that he couldn’t lift it?” Well, no. That would be a contradiction. Other constraints imposed by consistency may be more subtle. Like, why does God permit human history to proceed in certain ways, especially ways that we would much prefer that they didn’t? Here again, self-consistency probably plays an important role. Human free will is an important constraint. And there are likely other, unknown constraints, resulting from God’s unrevealed purposes.
Omniscience is the quality of knowing everything. This is also very related to being metaphysically absolute, the source of all things. As the cause of all things God also has knowledge of all things. If we imagine all things that can be known as a book God knows all things in that book, not only because he has read it, but also because he wrote it. He is the author of all that is. Many of the foregoing points about omnipotence apply here as well. There’s a classic concern about the conflict between divine omniscience and human free will. If God knows everything, including everything that we will ever do, can we really be said to freely choose to do those things? That’s a complicated problem and a whole topic in itself. Without actually resolving that question I’ll just make an observation using the analogy of the author. There is a sense in which the author of a story is constrained by the story itself. Authors can arbitrarily impose nonsensical decisions on their characters. But good authors don’t. Good authors follow their stories where they naturally lead. Their characters, even though they’re fictional, have a kind of free will of their own. That’s just an analogy but I think something similar applies to God’s authorship of all things and his knowledge of them. On the one hand he is the author and cause of all things. But this authorship and resulting knowledge is not just arbitrary. The evolution of all things, especially of human history, make sense and have a narrative coherence to them.
Finally, God is perfectly good. In Plato’s Republic, Socrates actually placed “the form of the Good” at the highest point on his spectrum of entities, the Divided Line. Goodness is not incidental to God’s nature but is absolutely intrinsic to who he is. One of the oldest problems in moral philosophy is whether God decrees what is good because it is good or whether it is good because he decrees it. This is a form of the Euthyphro Dilemma, based on another of Plato’s dialogues. Put another way, the question is whether God is prior to goodness or goodness prior to God. But in classical theism this is a false dilemma. God and the Good are not distinct at all. God is the Good.
Apart from classical theism the great worry with the Euthyphro Dilemma is that if goodness is merely whatever God decrees it to be then God could decree horrendous evils to be good. And they would have to be good. But under classical theism this is not possible. God is the Good. Neither God nor the Good are arbitrary. Horrendous evil cannot be made good and God cannot and will not decree them so. To do so would be to contradict his own nature.
All of the foregoing is principally philosophical rather than scriptural or based on revelatory religious experience. Though it has been most developed by Christians the foundations come largely from Platonist and Neo-Platonist philosophy, for example from Plotinus’s Enneads and Proclus’s Elements of Theology. Whether that is a weakness or a strength is a matter of perspective. I think it’s a strength but it also means that for Christian theology classical theism is a starting point rather than an end point. But I also consider it a great strength to see that classical theism spans so many traditions and schools of thought.
One of the best modern books on classical theism is David Bentley Hart’s The Experience of God. In that book he makes the following point:
“Certainly the definition of God I offer below is one that, allowing for a number of largely accidental variations, can be found in Judaism, Christianity, Islam, Vedantic and Bhaktic Hinduism, Sikhism, various late antique paganisms, and so forth (it even applies in many respects to various Mahayana formulations of, say, the Buddha Consciousness or the Buddha Nature, or even to the earliest Buddhist conception of the Unconditioned, or to certain aspects of the Tao…” (p. 4)
I find the Hindu convergences especially fascinating. Shankara (circa 700 – 750) was an interpreter of Vedantic Hinduism, Advaita Vedanta to be specific. A central concept in that tradition is Brahman, the highest universal principle, the ultimate reality, the cause of all that exists. In Advaita Vedanta this is identical to the substance of Atman, the Self or self-existent essence of individuals. Ramanuja (1017 – 1137) had a different interpretation called “qualified non-dualism” which makes greater distinction between Atman and Brahman. But Brahman, the ultimate reality behind all that exists, is central to the thought of both.
There are four modern authors on classical theism that I really like. These are David Bentley Hart, Edward Feser, James Dolezal, and Matthew Barrett.
I already mentioned David Bentley Hart’s book The Experience of God: Being, Consciousness, Bliss. Hart is an Orthodox Christian and also has an interesting affinity for Hinduism. In fact, the subtitle to his book – “Being, Consciousness, Bliss” – is a nod to the Hindu concept of Satcitananda, a Sanskrit term for the subjective experience of Brahman, the ultimate unchanging reality. Satcitananda is a compound word consisting of “sat”, “chit”, and “ananda”: being, consciousness, and bliss. These three are considered inseparable from Brahman.
Edward Feser’s book Five Proofs for the Existence of God goes through five proofs that he reworks from the ideas of five individuals: Aristotle, Plotinus, Augustine, Aquinas, and Leibniz. Each of the five proofs is classically theistic in nature. Later chapters in the book also go over the classical theist understanding of God’s nature in great detail.
James Dolezal’s major book on this subject is All That Is in God: Evangelical Theology and the Challenge of Classical Christian Theism. Dolezal pushes back on what he perceives as some drift away from classical theism in Evangelical theology. I mentioned earlier that some theologians place simplicity foremost among God’s attributes. Dolezal is one of these. Simplicity is central to his thought.
Matthew Barrett is a delightful theologian to read. He is editor of Credo Magazine and host of the Credo podcast. One of his common themes on Twitter is the need for Protestants and especially Evangelicals to take seriously the thought of Aquinas, the Church Fathers, and classical theism. His major book on the subject is None Greater: The Undomesticated Attributes of God.
Why talk about classical theism? To lay all my cards on the table, I desire for all to believe in God the Father, his Son Jesus Christ, and in the Holy Spirit. I am enthusiastically Christian and desire for all to be so as well, because I believe it is true. One of the first steps in this direction is belief in God. But in modernity belief in God is hardly a given. It might even seem implausible. How is believing in God any different from believing in Santa Claus, the Tooth Fairy, or the Flying Spaghetti Monster? Well, it’s actually extremely different. And I think that to really understand classical theism is to understand this difference.
God is not just an invisible being that we have to believe in, just because. Blind faith. Classical theism is much more philosophically reflective than that. To think about God is to think about and have some interest and curiosity about everything that exists, why it exists, and why it is as it is. It is maximally inquisitive and critically so. I believe that classical theism is very plausible and reasonable. That’s not actually why I believe in God or in Christianity. I attribute my belief to revelation from the Spirit. But intellectual openness and receptivity preceded that Spiritual revelation. Seeing classical theism to be a plausible and reasonable way to understand reality broke down intellectual and cultural barriers to spiritual receptivity. And that’s why I think it’s a topic worth talking about.